

I. Identification
Liomys salvini (Heteromyidae, Rodentia, Mammalia), or spiny pocket mouse, is the only member of the mouse family Heteromyidae that occurs in the ACG lowland dry forest (and in lowland dry forest elsewhere in Costa Rica, for that matter). The three other heteromyids in Costa Rica are the long and well known Heteromys desmarestianus of lowland ACG and Costa Rican rain forest (Fleming 1983), Heteromys oresterus of a few high elevation sites in the Cordillera de Talamanca, and the newly described Heteromys nubicolens known from just three localities (Monteverde, Rincon de la Vieja, y Volcan Cacao; Anderson and Timm 2006), rendering it endemic to the Cordillera Guanacaste and Cordillera Tilaran. Parenthetically, the ACG populations, and therefore this new species, of high elevation Heteromys was discovered by Marian Klaus when she was conducting a rodent inventory of the rain and cloud forests of ACG in 1998-1999. Adults of all ACG Heteromys are much larger than Liomys salvini, weighing twice or more as much, so they will not be confused with the dry forest Liomys. The two genera undoubtedly can be captured in the same ACG forest intergrade between dry forest and rain forest, or at least could be before most of it was deforested.
Heteromyidae, and therefore Liomys, can be distinguished from any other small ACG dry forest rodent (e.g., Sigmodon hispidus, Ototylomys phyllotis, Oryzomys palustris, Oryzomys fulvescens, Oryzomys alfaroi complex, Reithrodontomys, Nyctomys sumichrasti) by having fur-lined cheek pouches. The pouches are used for carrying seeds, insect pupae, and at times their own very young offspring, and evolutionarily formed by in-pocketing of outer body skin, which is the origin of the hair. Liomys salvini is brownish gray on its back and upper sides, and cream white below, while Heteromys desmarestianus and Heteromys nubicolens are gray on the back and sides, and white underneath, but with black forelegs. The face of Liomys is brownish gray with black eyes, while the face of Heteromys is grayish black with a light ring bordering the black eyes.
II. Geographic and microgeographic distribution
International
Liomys salvini ranges from the dry Pacific coastal lowlands of Oaxaca, Mexico to Costa Rica's dry forests (Fleming 1983). The northern margin of its distribution adjoins the southern distributions of three other species of Liomys (Liomys pictus, Liomys irroratus, and Liomys spectabilis) that occupy Guatemala and Mexico and even as far north as Texas (Brown and Genoways 1993).
Costa Rica
Liomys salvini occurs or ocurred in all Costa Rican dry forests below about 1200-1500 m elevation (Fleming 1983). Populations have survived at low and irregular densities in early secondary succession and on wooded margins of roads and pastures in many parts of Guanacaste Province and Puntarenas Province that were once covered with dry forest (and presumably had a continuous population of L. salvini). L. salvini is/was also present in some of the small pockets of relatively intact dry forest that remain in Costa Rica, such as those that were scattered over the area of today’s Area de Conservacion Guanacaste at the time of decreeing Santa Rosa National Park in 1971 (today’s Sector Santa Rosa).
Area de Conservacion Guanacaste
Today Liomys salvini occurs in all ACG dry forest of all ages, from the margins of the coastal mangroves to the lower slopes of the volcanos (300-1000 m). However, the upper elevation distribution may well be artificially generated by the massive deforestation of some of these slopes, which in turn dries and warms them, allowing lowland dry forest species to extend their populations upwards (i.e., as occurs with coyotes, white-tailed deer, ctenosaurs and cotton rats (Sigmodon hispidus)). The regenerating forest is richer in dry forest species of plants than was the evergreen forest that originally covered the sites. It appears that dry and deciduous forest is more friendly to Liomys salvini than is evergreen wet forest, though the detailed boundary and its intersection with Heteromys demarestianus and Heteromys nubicola populations has never been documented. When an old pasture (generally not occupied or used by Liomys) is regenerating its dry forest, it takes at least 20 years until the trees are old enough to be producing any serious quantity of the species of seeds that are desired by Liomys.
L. salvini is generally absent from pastures and other areas covered with herbaceous plants (which is to say, areas that produce quite small seeds and have little or no seed bank in the litter), except in years when there is a superabundant peak in the population (Fig. 4, 5, and see Janzen 1986). In these years there are L. salvini crossing all kinds of dryland habitats in their desperate search to locate habitat that is not already "owned" by other L. salvini.
L. salvini does not occur on the Islas Murciélago at the extreme western end of the ACG. This may be due to its elimination by predation by the introduced rats (Rattus norvegicus) which thoroughly occupy the islands. On the other hand, it could be that these islands do not have sufficient area (that to say, seed and insect pupae resources) to maintain a population of L. salvini. Certainly the very large grassy and herbaceous areas of the islands, due to the previous annual fires, are not suitable habitat for L. salvini. Today the islands are very impoverished in those woody plant species that produce seeds that are quality food for L. salvini, and I suspect that the elimination of Rattus would not result in a self-sustaining population of L. salvini being able to survive on the islands.
III. Natural history
Liomys salvini is a specialist at searching for, caching, and eating mature seeds of woody plants (and some herbs such as forest grasses) in ACG dry forest. IThe mice supplement this diet with moth pupae (and perhaps other dormant insects) found in the litter in both the rainy season and dry season. Seeds of the introduced/naturalized guanacaste tree (Enterolobium cyclocarpum; Fig. 3) offer a specific example (Janzen 1982a, 1982e, 1986). When the (re-introduced from Europe) range horses eat guanacaste fruits (as they would have in the Pleistocene, Janzen and Martin 1982), which they do very eagerly, the ungerminated seeds take 1 week to 6 months to pass through the horse (Janzen 1981a,b). These seeds are eagerly and thoroughly collected from horse dung by the mice, and more reluctantly from cattle dung (Janzen 1982d, 1986).

This individual is leafless at this time in the dry season, but has a very large crop of nearly mature fruits which darken the upper branch tips in this photograph. These fruits are nearly ready to fall to the ground and be eaten by the megafauna that would originally have consumed them and been seed dispersers (Janzen 1986, Janzen and Martin 1982). In ACG forest with few megafauna present (tapirs, peccaries, horses and cattle in the old days),the seeds are thoroughly harvested and preyed upon by Liomys salvini, either by gnawing them out of the fruits or by harvesting them from the dung of seed dispersers (March 1972, Bebidero, Guanacaste, Costa Rica).

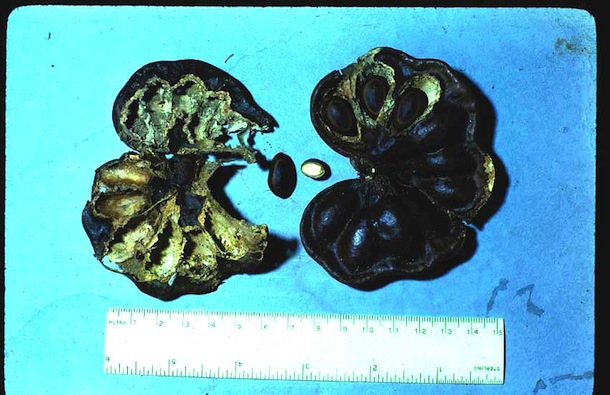


In ACG dry forest, in the evening after dark, the Liomys leave their below-ground tunnels where they nest and store food, and search for seeds in the litter, in fallen fruits, and in the dung of other animals (currasows, coatimundis, horses, ctenosaurs, etc., Janzen 1986). The seeds that are acceptable are stuffed into their cheek pouches with their front legs, and carried to their underground storage rooms (small rooms 0.2 to 1 liter in volume off the sides of their tunnels) to be eaten later. Some species of seeds are eaten at the moment of encountering them or stored, depending on how hungry the mouse. Other species of seeds are rejected because of the presence of defensive chemicals (Fig. 8). No seed is so hard that the mice cannot open them (however, the fruits of guapinol, Hymenaea courbaril, are so hard and large in diameter that L. salvini cannot get to their seeds unless some other animal such as an agouti or tepiscuintle opens them) (Fig. 1).
In addition to seeds, the mice search for and eat moth pupae, and sometimes insects, that they find in the litter and upper soil layer. There is no evidence that they climb trees for any reason, but in human habitations they learn to climb on furniture and counter tops in search of food.
L. salvini searches for seeds in the litter of the forest understory at any time during the night (not in the daytime), but every hour spent searching in the litter is another hour exposed to various predators. The mice are very disinclined to search for seeds when there is a full moon (lighting the forest understory), and when the litter is very dry, which makes search for seeds very noisy. It is likely that a given mouse leaves its tunnels to search in the litter every 1-3 nights. When it finds a large source of seeds, such as a newly fallen fruit or seed crop in the litter, it can spend many hours transporting seeds from the source to its tunnels.
A mouse may, for example, eat 3-7 newly germinated guanacaste tree seeds per night. While a mouse may survive for months in the laboratory on a pure diet of certain species of seeds (for example, Sesbania emercus, Guazuma ulmifolia, Enterolobium cyclocarpum, Apeiba tibourbou, Malvaviscus arboreus, Cochlospermum vitifolium, Bursera simaruba), in nature a mouse's diet is much more mixed (Janzen 1982a,b,c, 1986). It is quite noticeable that in captivity L. salvini will die of hunger before eating the species of seeds that it rejects in the forest (e.g., Pithecellobium platylobum, Lonchocarpus felipei (Fig. 8), Swietenia macrophylla) (Janzen et al 1990). Enterolobium cyclocarpum seeds are a much better diet when germinated than when hard (dormant), and the mice cut the hard seed coat (Fig. 7) and leave the seeds on moist soil in their tunnels to germinate (Fig. 7) before eating them. They are, in effect, making bean sprouts.
Outstanding physical traits
Liomys salvini weighs 7-13 g when it leaves its mother's nest (fledges) or is expelled by her, and grows to weigh 35-45 g (female; 55 g when very pregnant) or 45-75 g (male). While this mouse normally walks on all four feet, when it is in a hurry or is startled it runs by jumps and may jump as far as 1.5 m in one leap. It is likely that this ability to jump is the antecedent for the great jumping ability of its evolutionary descendent, the kangaroo rat (Dipodomys spp.) of northern Mexico and the southwestern USA. Dipodomys is evolutionarily derived from a heteromyid such as Liomys (Genoways and Brown 1993).
The hairs on the back of Liomys salvini are of two kinds: thin and "normal", or thickened into a flat weak spine, as if they were evolving in the direction of the ACG’s prehensile-tailed porcupine’s spines (Sphigurrus mexicanus). These "protospines" seem to make the mice much more slippery and hard to hold on to than if they were normal hair, because the approaching hand (or predator) cannot pass them to grab the skin and body. Their stiff sharp points also probably resist an approaching mouth or claw.
Reproduction
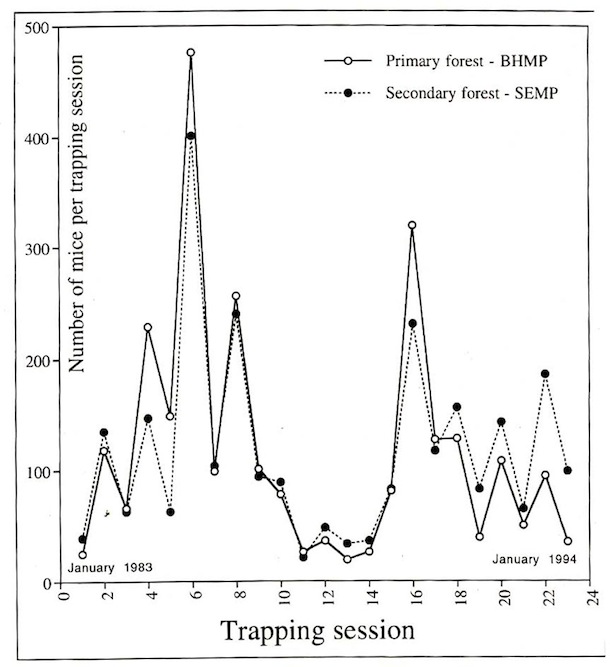
When the population density of Liomys salvini is moderate to high (50-400 mice per 4.5 hectares of forest study plot), the general pattern is that the adult females give birth to a 3-4 member litter between mid-January and mid-February (i.e., the first two months of the dry season, and the dry season is the time of the maximum number of seeds in the litter, both because they are not escaping the mice by germinating and many species of dry forest trees mature their fruits in the dry season (Janzen 1967). These are weaned and out of the nest (whether pushed by the mother or gone on their own is not known, but it is probably the former) by March-April. Adult females often produce a second litter in May-June (beginning of the rainy season). During the remainder of the rainy season (when seed density is rapidly declining owing to germination and trees being non-reproductive), the males have shrunken epididymis and only partly descended testes, and pregnant and lactating females seem to be almost entirely absent. However, when the density of Liomys is very low (e.g., 1-25 mice per 4.5 hectare forest study plot), there are some reproducing females in the rainy season, as evidenced by 3-5 month old mice captured in January. This pulsed, episodic reproductive behavior results in the “average” female usually not reproducing in the first calendar year (February-December) of her life, then 1 or 2 times in the second year, and (if she lives this long) again 1-2 times in her third calendar year. For males, the pattern is roughly the same. Just as only some of the “young of the year” females reproduce for their first time in May-June, and the same for males, Liomys are both not in continuous reproduction and must survive year-long-periods between birth and insemination events; this is very different from the other small rodents in ACG dry forest, which appear to reproduce throughout the year. I feel that one of the consequences of this reproductive pattern is that Liomys individuals are more predator-wary than are other species of small rodents, leading among other things to both strong territoriality and a more detailed and intimate knowledge of the specific details of that territory (multiple nest entrances, obstacles, distances, etc.). From a practical census standpoint, this means that by trapping the mice in January they are caught at their lowest density and in May they are at their highest density (Fig. 15), with the numbers gradually falling from June through December as predators take their toll.
Population
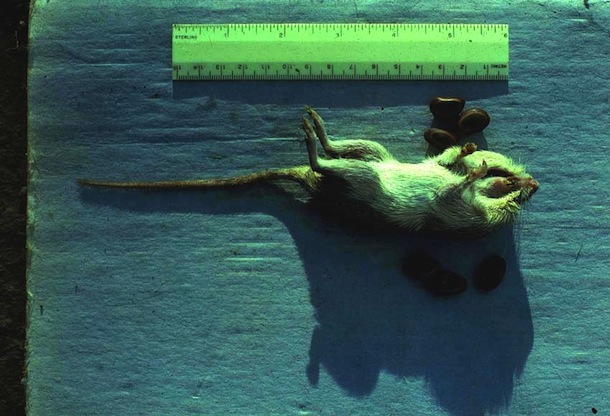
Through capturing Liomys salvini with Sherman live traps (Fig. 9, 10, 11), marking them with tattoos (Fig. 12,13), and releasing them for recapture later, it has been possible to monitor their density and individual locations in two 4.5 ha plots for 30 years in the Bosque Humedo and Bosque San Emilio, Sector Santa Rosa (Fig. 14). The results (Fig. 15) show clearly that the density of L. salvini fluctuated very strongly and with a peak-to-peak cycle of 4-5 years in the beginning, but then leveled off at a very low density (but notably not extinguished) since then.
These population fluctuations during the first 10 years of monitoring provokes many thoughts as to how it would affect both the many animals that eat Liomys and the many seeds that Liomys eats, to say nothing of the ease of noticing and studying these mice.



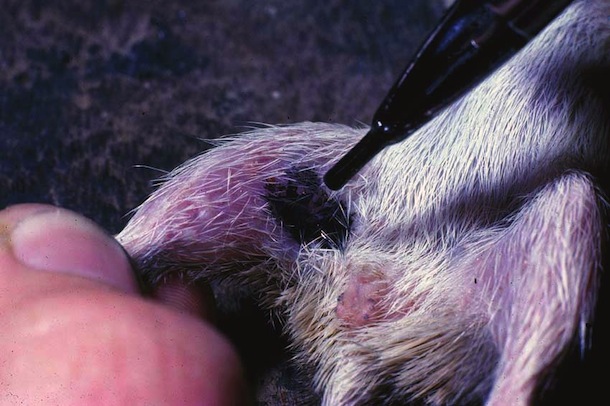
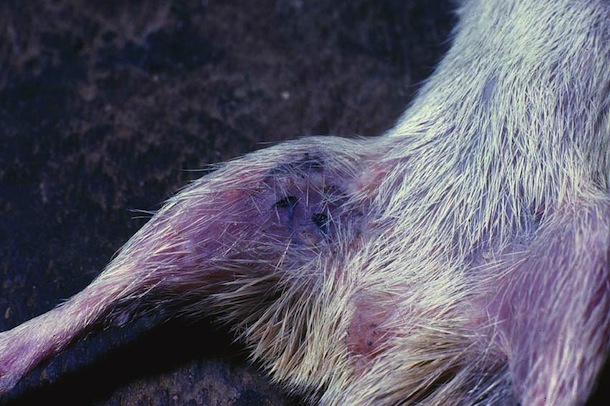

Parasites and Predators
Liomys salvini is prey for many species of nocturnal predaceous vertebrates (snakes (Fig. 16-17), cats, owls, skunks, possums, coyotes, gray fox) that live in ACG dry forest. It seems that the ups and downs in the population in Fig. 15 are generated by the interaction between the mice and these predators. When the density is very low, the predators lack one of their principal foods and generally do not breed, and their density falls. Then, the mouse population grows very rapidly, thereby generating lots of food for the predators, which in turn have a bout of reproduction. This increased population of predators then lowers the mouse population again. In the 1983-1987 peak this process was cleanly executed, while in the 1990-1994 boom-bust cycle the relationship was less clear, probably due to the six successive years of excessively low rainfall that occurred during this period.
Based on early survey of their parasites, it is known that Liomys salvini in the ACG supports a rich fauna of mostly undescribed tapeworms, nematodes, moths (Davis et al 1986), pseudoscorpions, lice, mites and ticks. It does not, however, have any fleas.


IV. How to find a Liomys salvini
When the density of Liomys is very low, as it has been since 1996-1997 there is no reliable way to “find” a Liomys other than setting a very large number of Sherman live traps. When the density is high, setting as few as 10 Sherman live traps in Sector Santa Rosa dry forest more than 50 years old will usually catch a Liomys during the first night of trapping. When densities are high, it is also relatively easy to meet a free-foraging Liomys in the forest at night by just walking quietly in the forest and searching with a headlamp. Try to catch it by hand and you will discover how incredibly rapidly a Liomys can avoid a predator. A relatively easy way to see one is to ask about the detailed live-trapping schedule in January or May, and it can be arranged for you to meet at least a few of them (to meet both sexes may require several nights of our trapping in Bosque San Emilio). Alternatively, even with very low density, we occasionally have 1-2 of them foraging on the floor of our very open house in Area Administrativa of Sector Santa Elena, and we can tell you by e-mail if there is a regular visitor at any given time.
Literature cited
Brown, J. H. and Genoways, H. H.,eds. 1993. The biology of the Heteromyidae. Special Publication No. 10, The American Society of Mammalogists, 715 pp.
Davis, D. R., Clayton, D. H., Janzen, D. H., and Brooke, A. P. 1986. Neotropical Tineidae, II: biological notes and descriptions of two new moths phoretic on spiny pocket mice in Costa Rica (Lepidoptera: Tineoidea). Proceedings of the Entomological Society of Washington 88:98-109.
Fleming, T. 1983. Heteromys desmarestianus (Raton Semiespinosa, Spiny Pocket Mouse). Costa Rican Natural History, pp. 474-475, ed. D. H. Janzen, University of Chicago Press, Chicago.
Janzen, D. H. 1967. Synchronization of sexual reproduction of trees with the dry season in Central America. Evolution 21:620-637.
Janzen, D. H. 1981a. Enterolobium cyclocarpum seed passage rate and survival in horses, Costa Rican Pleistocene seed dispersal agents. Ecology 62:593-601.
Janzen, D. H. 1981b. Guanacaste tree seed-swallowing by Costa Rican range horses. Ecology 62:587-592.
Janzen, D. H. 1982a. Seed removal from fallen guanacaste fruits (Enterolobiumcyclocarpum) by spiny pocket mice (Liomyssalvini). Brenesia 19/20:425-429.
Janzen, D. H. 1982b. Fruit traits, and seed consumption by rodents, of Crescentiaalata (Bignoniaceae) in Santa Rosa National Park, Costa Rica. American Journal of Botany 69:1258-1268.
Janzen, D. H. 1982c. Natural history of guacimo fruits (Sterculiaceae: Guazuma ulmifolia) with respect to consumption by large mammals. American Journal of Botany 69:1240-1250.
Janzen, D. H. 1982d. Attraction of Liomys mice to horse dung and the extinction of this response. Animal Behaviour 30:483-489.
Janzen, D. H. 1982e. Removal of seeds from horse dung by tropical rodents: influence of habitat and amount of dung. Ecology 63:1887-1900
Janzen, D. H. 1986. Mice, big mammals, and seeds: it matters who defecates what where. In Frugivores and seed dispersal, A. Estrada and T. H. Fleming, eds., Dr. W. Junk Publishers, Dordrecht, Holland, pp. 251-271.
Janzen, D. H., Fellows, L. E., and Waterman, P. G. 1990. What protects Lonchocarpus (Leguminosae) seeds in a Costa Rican dry forest? Biotropica 22:272-285.
Janzen, D. H. and P. S. Martin 1982. Neotropical anachronisms: the fruits the gomphotheres ate. Science 215:19-27.